
The plant micro-biome, a tool to help plants survive climate change?
- Michael Remke, Nancy Johnson, Matthew Bowker
- Sep 26, 2017
- 9 min read
Many areas on the Colorado Plateau are experiencing extreme drought which coincide with warmer temperatures, more frequent fires, and insect outbreaks. These factors are resulting in rapid vegetation change including widespread mortality of several dominant plant species. The result is often landscapes scared with erosion issues as they we lose the services that living vegetation provide. Many efforts to restore these landscapes often show little success, especially in drylands.

Up to 90% mortality of Engelmann Spruce has been observed at scales of thousands of acres in the San Juan Mountains, Colorado. This photo represents these data and provides context to the landscapes where this is occurring.
Plant restoration strategies following these disturbances historically have focused only on the reintroduction of native plant materials, however, we may be missing a hidden world that may facilitate plant success in restoration, soil. Soils are rich in complex biotic communities, many of which are analogous to a plant micro-biome. Microbes such as bacteria and fungi help plants gather soil water and nutrients through a variety of enzymatic and exploratory structures. One key group of organisms in this plant micro-biome are mycorrhizal fungi. Mycorrhizal come from myco, meaning fungus, and rhizo, meaning root, so they are literally root fungi. They live in or on plant roots and develop extensive networks of hyphae, think tiny filaments, that can efficiently gather water and nutrients from the soil. These fungi then trade the nutrients and water they gather for carbon from the plant. What if then, we used these fungi in restoration to help improve plant performance?

These Golden Chanterelles are actually mycorrhizal to pine trees and firs, forming up 100 meters of fungal hyphae in a single gram of soil.
Studies conducted on this topic have been done in greenhouses and have found limited application in field settings, to the contrary, field studies focused on soil biota show success in accelerating restoration goals. I propose that symbiotic soil organisms may be the critical component for enhancing success of plant restoration trials, especially in the context of climate change. Because of how efficient mycorrhizal fungi are in gathering soil water, perhaps they may help plants survive drought. My project has already and will continue to enhance the scientific community’s understanding of mechanisms that may preserve plant productivity in the face of a changing climate while also exploring a novel tool in plant restoration. If this proves to be successful, we will also be able to preserve ecosystem function and the ecosystems services to which human societies are accustomed. This clearly benefits science by filling gaps in scientist’s knowledge and humanity by finding mechanisms that preserve ecosystem services.

Infrastructure in place for a field experiment. Moving away from greenhouse studies can help us better understand the complex ecology and plants and their associated microbes in a more variable setting.
I continue to work with two plant species – one herbaceous plant, Bouteloua gracilis (Blue Grama) and one tree, Pinus ponderosa (Ponderosa Pine) – in a field manipulation experiment. Each of these species forms different relationships with different types of soil organisms. This experiment uses one focal site for each plant species and several transplant sites along an elevation gradient to induce both warming and cooling scenarios. I manipulated four factors: 1) Background soil origin – Soil types change over small geographic space, so each plant will be grown in sterilized soil from either their site of origin or the site they are transplanted to. In addition, plants will be grown in a sterile nursery potting soil to simulate the action of land managers. Land managers rarely inoculate plants with soil organism communities prior to field planting. 2) Soil organism origin – Each plant will be grown with live soil inoculum from either their site of origin or their transplant site. A one-centimeter thick layer of field soil has been shown sufficient to inoculate plant roots with symbiotic soil organisms, thus making it an easy factor to manipulate. 3) Temperature – using an elevation gradient will create a temperature gradient. By moving plants from their site of origin to sites of lower elevations, I will be able to simulate warming. Plants will be transplanted to sites that are approximately 2ºC and 4ºC warmer. For each transplant, some plants will be transplanted back to their site of origin as a control. Similar transplants will be made to sites of higher elevation to simulate assisted migration. 4) Moisture – Since precipitation generally decreases with elevation, each of the plants being transplanted downward to simulate warming will be grown under either ambient conditions of the transplant site or with supplemental water. This is an important distinction to make since not all climate models are in agreement as to how precipitation will change in the future.

As soil types change in the same environment, so do plant communities. Here, a basalt lava flowed over the Moenkopi sandstone, in the image the red sand stone hosts a less diverse plant community with lower plant cover. By manipulating in soils in experiments, we can better learn about how they influence plants.
When I completed these manipulations, I was then able to test several hypotheses related to vegetation change and assisted migration. I continue to determine if assisted migration is possible simply by transplanting plants to new sites, or whether the plants need their “home site” soil organisms for success. As I manipulate both the background soil environment and soil organisms I will determine if success of assisted migration is likely to be limited due to changes in soil types across regions. I am also testing how well plants can adapt to climate change in situ and if pairing plants with the proper soil organism community gives them an advantage.
I am illuminating some of the basic rules dictating the degree of mutualistic function in plant-soil organism interactions, with this process. Since each of these plants forms associations with unique combinations of soil organisms, they each may respond differently. For example, some sites may limit the establishment of certain plants if their associated soil organisms are missing. Lastly, by growing plants in a standard nursery potting soil mix and pairing them with soil organisms, I am able to quantify the benefits of adding soil organism treatments to standard nursery practices. It may be the case that plants are dependent on soil organisms from their home site to adapt to novel conditions.
I now have three years of data with blue grama in the field and the emerging patterns are quite compelling. The most striking result thus far is that plants consistently grow larger when paired with their “home team” soil organisms. This is especially true for plants growing in warmer, drier environments. Interestingly, the size of this effect starts to diminish in cooler, more moist environments, and in fact no longer makes a difference. Another important result thus far is that mortality has begun to occur in plants planted at sites that are two and three degrees warmer than the site they came from, only if, they are not growing with their “home team” soil organisms, and the rate of mortality is exacerbated when plants are also growing in a unique soil type. These results show the striking importance of soil biota in facilitating plant growth, especially in drought like scenarios or stressful environments that could be created by phenomena like climate change. Thus, it is imperative that land managers consider soil disturbance and we begin exploring economically viable ways to restore soil biota to enhance our plant restoration efforts.
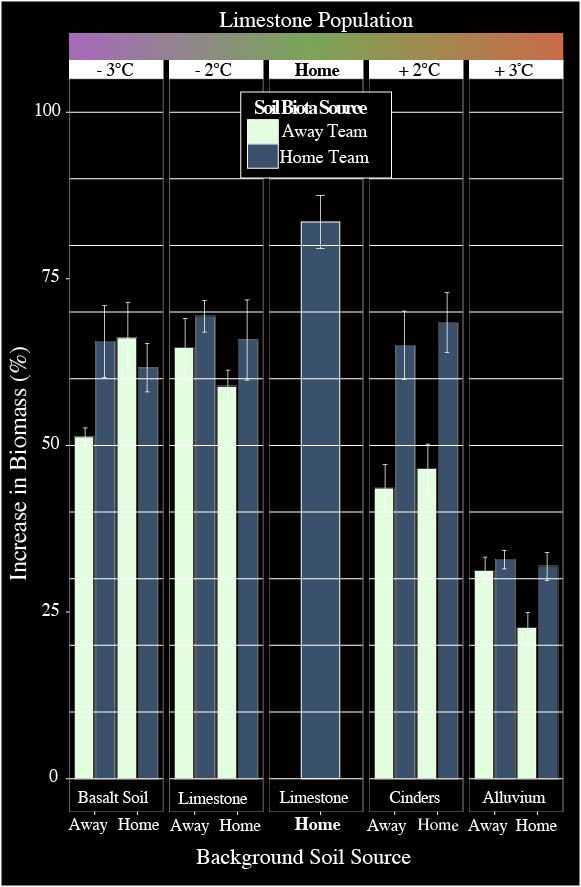
This graph represents blue grama growth since the time of planting. On the Y-axis is increase in plant biomass since time of planting. A 100% increase would be a doubling in size. Along the X-axis is the different sites with different soil types. Dark colors represent home team parings, light colors represent away team.


Above, blue grama grown with the home team at the site that is 2C warmer contrasted to below, blue grama grown with the away team.
I have now completed one year with Ponderosa pine in the field. Based on data from blue grama, it is not surprising that Ponderosa pine also grew larger when we paired it with its “home team” soil organisms in a warm and dry setting relative to Ponderosa pine’s site of origin. In fact, trees planted at the warm dry site with their home team soil organisms grew nearly 50% more than ponderosa pine planted at its site of origin. This result is backed by metrics of plant physiology that suggest in the warm, dry environment, the pines without their “home team” soil organisms close their stomata to prevent water loss, thus inhibiting photosynthesis. Physiology data also suggests these trees have access to more water than trees grown without their home team soil biota, making a compelling argument that the home team benefit is a story of moisture. The trees are obtaining more water and not becoming water stressed despite receiving the same amount of precipitation. This line of evidence adds compelling depth to the conversation because it demonstrated that the “home team” soil organisms are helping the trees acquire water and maintain higher photosynthetic rates thus facilitating greater growth. Perhaps this moisture story combined with warm springs and longer growing seasons at the warm site relative to the site of origin explains why these trees could grow so much more in a warmer world.

This graph represents ponderosa pine growth since the time of planting. On the Y-axis is increase in plant biomass since time of planting. A 100% increase would be a doubling in size. Along the X-axis is the different sites with different soil types. Dark colors represent home team parings, light colors represent away team.


Above, blue grama grown with the home team at the site that is 2C warmer contrasted to below, blue grama grown with the away team.
Unfortunately, the majority of the trees planted at the cool wet site died this spring. We had a very wet winter and the geomorphology of this site creates a shallow water table that had all our seedlings roots saturated in a vicious freeze thaw cycle all spring. Since we lost most of these trees we really cannot speak to why or how they died but it does bring a word of caution with regards to assisted migration, or any artificial planting project. Site conditions matter. Just because there are mature trees does not mean young trees can establish on site today in the given climate context. Also, geomorphic and soil conditions interact with climate to create site conditions. In our case a deep clay layer restricted water flow through the soil profile resulting in the saturated environment we observed following snowmelt, this setting would likely cause any artificial planting to fail.
Perhaps the biggest benefit of this project is the multitude of environmental contexts in which we are studying these interactions. I don’t mean to suggest that these data tell us soil biota are always beneficial, but rather am hoping to build their importance into the contexts of dryland ecosystems. In wetter environments, an abundance of scientific literature shows how soil pathogens accumulate to attack their host plants and thus escaping their soil biota is beneficial. This is known as the enemy release hypothesis and is likely an important mechanism in maintaining. diversity in very mesic, diverse systems – like tropical rainforests. In contrasts, dryland systems have low diversity and we suggest soil organisms If we are going to spend money on intentionally manipulating ecosystems and planting trees, we better ensure we are doing it in a constructive way. Overall, these findings show the crucial importance of soil biota in facilitating plant growth and survival in a variety of different environmental conditions across the dry ecosystems of the Colorado Plateau.

The tropics of Belize are clearly a much more diverse plant community and likely have very different associations with soil biota. Here a battle with pathogens dominates the story, rather than a tale of mutualists. Its important to understand the environmental context of the system you are working in.
This perspective needs to be transferred to more applied projects as we continue to monitor and expand this project over the years. We hope to invest time into understanding how we could create seed coats with native soil biota, thus inoculating plants at the time of planting with their appropriate micro-biome. We are currently working with the Czech Academy of Sciences to assess the community of these plants micro-biomes and how these communities change over time. We hope this will answer several questions in regards to what species are doing what and how they interact with different plant populations, or if the same species behaves uniquely from different populations and environments. I hope you will continue to follow our work as we explore how the plant micro-biome interacts with plant populations and changes in the environment. Let’s bring back the invaluable ecosystem services of the Colorado Plateau.

The drylands of the Grand Canyon make a perfect location to imagine the role of these soil biota, and I just wanted to leave you with a pretty picture of the desert. Thanks for reading.
Special thanks to the Grand Canyon Trust, McIntire Stienis Foundation, NAU School of Forestry Czech Ministry of Sciences, Babbit Ranches, Bureau of Land Management, Colorado Plateau Native Plant Program, United States Forest Service, the Bowker and Johnson Lab groups and the dozens of hands that have helped with the arduous mental and physicall work put into this project.



Comments